

 分享
分享

哮喘作为一种复杂的高度异质性疾病,其临床表现涵盖了多样的症状和复杂的表型。此类表型的形成由细胞与分子通路等复杂生物学机制驱动,而随着对哮喘病理机制的认知不断深化,生物靶向治疗已成为重度哮喘临床治疗的核心手段,该疗法通过精准作用于特定炎症通路或分子靶点,实现了重度哮喘管理模式的突破性优化。
传统靶点:下游抗炎的局限与困境
在炎症性疾病的治疗历程中,传统靶点如白细胞介素(IL)-4、IL-5、IgE等发挥了重要作用,为众多患者带来了临床获益。然而,这些传统靶点都处于炎症反应的下游环节。当过敏原入侵呼吸道,气道上皮细胞首先感知到危险信号,启动免疫反应,被激活的免疫细胞随即产生IL-4、IL-5、IgE等细胞因子,促进嗜酸性粒细胞的浸润,形成典型的2型免疫反应[1]。它们只能对已经产生的炎症反应进行干预,而无法源头阻断。从炎症网络的角度来看,它们的作用局限于单一或少数几条炎症路径。比如,IL-4主要诱导辅助性T细胞2(Th2)分化,IL-5促进嗜酸性粒细胞分化与成熟,而IgE主要介导过敏反应。这就导致当炎症网络反应中多条通路同时被激活,或者炎症机制发生改变时,仅针对下游靶点进行干预,无法从根本上切断炎症的根源,也难以全面抑制炎症的发展[2]。
上游信号:炎症网络的 “总开关”
气道上皮作为呼吸道的第一道防线,过敏原、病原体、空气污染物、化学物质等激活上皮细胞,导致上皮细胞活化并释放胸腺基质淋巴细胞生成素(TSLP)、IL-25和IL-33,进而激活Th2细胞、2型固有淋巴细胞(ILC2s)和树突状细胞,并导致这些细胞过度产生IL-4、IL-5和IL-13[3]。从炎症反应整体来看,这些预警素作为免疫应答的"启动信号",处于一个极为特殊且关键的位置,它就像是整个炎症级联反应的“启动按钮”,一旦被触发,便开启了一系列复杂而广泛的炎症反应,相较于传统的下游靶点,它的作用更具前瞻性和根本性。预警素的强大之处不仅在于其处于炎症反应的上游,更在于它能促进下游炎症细胞活化,释放炎症因子,激活多条炎症通路,具有调控能力。
TSLP主要由上皮细胞产生,成纤维细胞、树突状细胞、巨噬细胞及肥大细胞亦可少量分泌。在2型炎症反应中,TSLP发挥着核心驱动作用。该因子在机体接触过敏原、抗原、病毒及污染物后释放,可激活包括ILC2s在内的多种效应细胞,并促进此类细胞分泌IL-4、IL-5、IL-9及IL-13。其中,IL-4可诱导Th2分化并促进IgE的合成;IL-5 是嗜酸性粒细胞性炎症发生的关键驱动因子;IL-9 能够刺激肥大细胞增殖;IL-13 则会引发杯状细胞增生,同时造成气道平滑肌细胞高反应性与过度增殖[4]。
IL-33由多种支气管上皮细胞产生,在上皮损伤后迅速释放并持续存在,即使在过敏原暴露停止后仍可维持炎症活性。它可激活ILC2产生IL-13和IL-5等2型细胞因子,还可改变肥大细胞的表型与组织浸润,并导致杯状细胞黏液化生和气道堵塞[5]。
IL-25可诱导记忆性Th2活化与极化,促使其分泌IL-4、IL-5和IL-13。IL-25还可通过直接结合嗜酸性粒细胞表面的IL-17RB,在延缓嗜酸性粒细胞凋亡、维持过敏性气道炎症状态下嗜酸性粒细胞增多症病程中发挥关键作用[6]。
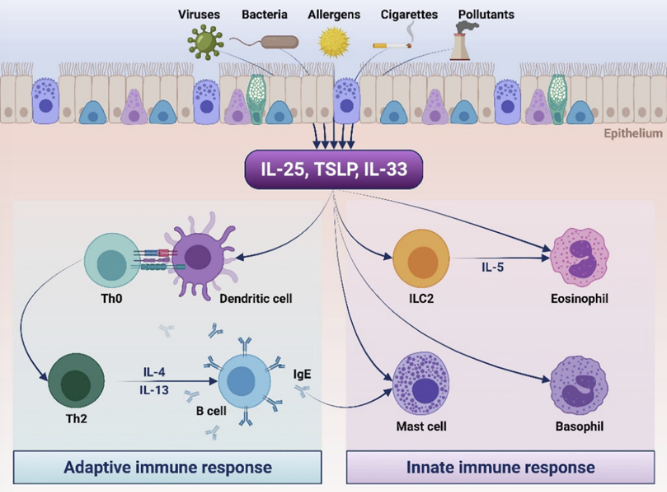
图1 哮喘中预警素的致病作用[4]
从源头阻断:炎症治疗的新曙光
针对TSLP、IL-33、IL-25等上皮源性预警素的靶向干预策略,最核心的优势在于实现炎症反应的“源头阻断”。该策略通过靶向抑制免疫调控网络上游的关键驱动分子,可广泛调控以Th2型免疫应答为代表的多条炎症通路,克服了传统下游靶点药物作用范围局限的短板。从作用机制分析,此类干预聚焦于炎症反应的起始环节,其作用模式类似于构建防火隔离带,能够从根源上阻断下游 IL-4、IL-5、IL-13等效应细胞因子的级联激活与信号放大,进而高效遏制炎症反应的全面爆发,同时有助于减少因单一通路阻断所诱发哮喘炎症类型转换风险[5]。与仅靶向阻断单一下游效应分子的治疗策略相比,针对炎症反应上游靶点的干预手段,可在炎症级联反应启动阶段同步调控多条下游信号通路。该策略在合并多因子共激活、多种生物标志物同步升高的混合炎症表型患者中,展现出潜在的应用优势,为突破现有治疗困境提供了全新的解决思路。
总结
重度哮喘具有高度异质性,与多种炎症细胞和炎症因子的异常产生和释放有关,现有生物制剂主要针对哮喘下游炎症因子,难以满足复杂的哮喘患者治疗需求。而TSLP、IL-25、IL-33 等上游预警素作为炎症“总开关”,位于各类炎症通路上游,启动和放大不同类型的炎症反应,未来围绕这类预警素的上游干预,有望成为突破哮喘精准诊疗瓶颈的关键方向。
参考文献
1. Bart N Lambrecht et al. Nat Immunol, 2025. 26(8): p. 1233-1245.9.
2. Lindsley AW, et al. J Asthma Allergy. 2025 Jan 14; 18: 33-57.
3. Haiyu Hong et al. Allergy. 2020 Nov;75(11):2794-2804.
4. Corrado Pelaia et al. Biomedicines. 2021 Aug 29;9(9):1108.
5. Ayobami Akenroye et al. J Allergy Clin Immunol. 2025 Apr;155(4):1133-1148.
6. Florence Roan et al. J Clin Invest. 2019 Apr 1;129(4):1441–1451.
7. Cheung PF et al. Allergy. 2006; 61(7): 878-885.
CN-177246,截止至2027-01-20
本材料由阿斯利康提供,仅供卫生专业人士参考,不用于推广目的
本文完
责编:Jerry